|
|
Results from a large number of studies indicate that variability in blood pressure is responsive to a variety of environmental, physiological, and sociocultural stimuli. For example, age is a strong predictor of blood pressure in most representative samples from more cosmopolitan societies, including African Americans, European Australians and European Londoners; however, age is little associated with blood pressure in groups still following more traditional lifeways, including the Yanomani, Gainj and Cofán [178, 179, 180, 181]. Conversely, associations of body mass index with blood pressure appear to be more similar across populations and environments [182].
Although numerous reasons have been advanced to explain population differences in blood pressure, including variations in psychosocial stressors, sodium intake, diet, ecological factors, and underlying predispositions associated with ethnicity and biology, no clear explanatory model for blood pressure differentials has yet emerged.
In Western communities high blood pressure affects 15-20% of the adult population and is a significant contributor to morbidity and mortality from cardiovascular disease including stroke, myocardial infarction and end-stage renal disease [14, 183]. Some preliminary investigations of genetic control of blood pressure have yielded a rough estimate of the proportion of trait variation attributable to genetic effects. Genetic heritability evinced in these studies has ranged from 0.14 to 0.82 for systolic blood pressure and from 0.08 to 0.64 for diastolic blood pressure, depending on age of the studied individuals [18, 184, 185, 186, 187].
In all human populations studied so far the distribution of both systolic and diastolic blood pressure follows an unimodal, continuous distribution. This distribution of blood pressure strongly supports the hypothesis that individual blood pressure is determined by a complex interaction of several susceptibility genes and the environment [188].
Identification of these genetic factors is of critical importance because it will permit a greater understanding of the pathogenic mechanisms of hypertension [189]. Since the risk of stroke, coronary artery disease, congestive heart failure and renal insufficiency increases with increasing blood pressure and operates over the whole range of blood pressure without threshold [190], the definition of susceptibility genes for hypertension may provide important information on genetic factors contributing to coronary artery disease and stroke. The potential of such genetic markers to predict individual risk of end-organ complications and to tailor therapies may facilitate accurate targeting of drug therapies and perhaps development of novel treatments.
Methods currently used to identify and locate genetic contributors to hypertension are based on association studies with marker loci, linkage studies, and candidate genes [42]. Because of their intimate involvement, two main physiological systems have borne the brunt of genetic studies on blood pressure regulation: the sympathetic nervous system and the renin-angiotensin system (RAS). The sympathetic nervous system influences cardiac output, vascular tone, renal sodium reabsorption, and is therefore closely involved in the regulation of blood pressure [141]. The sympathetic neurotransmitters, the catecholamines epinephrine and norepinephrine, mediate their effects on the various target organs by interaction with membrane-bound adrenergic receptors. Activation of α-adrenergic receptors causes vasoconstriction and a rise in blood pressure, whereas activation of β2 receptors by specific agonists leads to vasodilatation and a consecutive fall in blood pressure. Recently, the α1B and the dopamine 1A receptor have been linked to systolic blood pressure variations in young Caucasians [191]. The β2 adrenoceptor gene, which codes for a key component of the sympathetic nervous system, has been identified as a potential candidate gene for the development of high blood pressure [192, 193, 194]. For this reason, we focused on the β2 adrenergic receptor in this study.
Recently, significant genetic variability in the structure of the β2 adrenoceptor in the human population due to polymorphisms has been found (Table 5.1) [192, 195, 196]. These occur in the amino terminus of the receptor at amino acids 16 (Arg or Gly), 27 (Gln or Glu) and 34 (Val or Met) and in the fourth transmembrane spanning domain at amino acid 164 (Thr or Ile). In recombinant cell studies [197, 198], and in primary cultures of cells endogenously expressing these variants [199], clear phenotypic differences have been shown between the polymorphic receptors. The Gly16 receptor was found to undergo enhanced agonist-promoted downregulation of receptor number as compared with the Arg16 receptor [197]. In contrast, the Glu27 receptor was found to undergo very little agonist-promoted downregulation compared with the Gln27 receptor [197]. These variants are common in the population (Table 5.2). Met at position 34 is very rare (< 1% of the population), and it has been shown that this substitution does not alter receptor function [200]. The Ile164 receptor, which occurs in the heterozygous state in ~5% of the population, displays depressed coupling to the stimulatory G protein, Gs.
Cellular expression of the β2 adrenergic receptor is controlled in part by a 19-amino acid peptide that regulates mRNA translation. This peptide is encoded by a short open reading frame (sORF), termed 5’ leader cistron (5’ LC), which is 102 bp upstream of the β2 adrenoceptor coding block, referred to as BUP (β2 upstream protein). Mutation analysis of the 5’ leader region and peptide coding sequence suggests this peptide impedes translation of β2 adrenoceptor mRNA, and thus regulates cellular expression of the receptor [104]. A recently defined polymorphism at position 19 of this 5’ leader cistron (Cys19→Arg19) has been shown to reduce the effect of the BUP driven β2 receptor expression inhibition [171]. β2 Adrenoceptor density was 110% higher in cells transfected with the 5’ LC-Arg19 construct as compared with those transfected with the 5’ LC-Cys19 construct. Levels of the two mRNA transcripts were not different, confirming in vitro studies that the upstream peptide regulates receptor expression at the translational level. Polymorphisms of the 5’ LC are in linkage disequilibrium with polymorphisms at position 16 and 27 of the β2 adrenergic receptor gene, so that 5’ LC Arg19 occurs more frequently with Gly16 and Glu27.
The African Caribbean Study. In this study, we have demonstrated association of a variant within the β2 adrenoceptor gene with essential hypertension in African Caribbeans. The frequency of the prodownregulatory Gly16 allele of the β2 adrenoceptor gene is considerably higher in hypertensives than in normotensives. This observation may account for β2 adrenergic receptor regulation in resistance vessels and might explain blunted forearm vasodilatation that has been reported in normotensive African Americans in response to infusion of the β-adrenoceptor agonist isoprenaline [201]. Such attenuation of the vasodilatory response in people of West African ancestry could contribute to the increase in total peripheral resistance in response to sympathetic stimulation by mental or physical stress and may elevate blood pressure [145].
This result is very unlikely to be due to ethnic admixture or a population-stratification artifact, since the distribution of other diallelic polymorphisms and blood-group antigens within St. Vincent is very similar to that observed in West Africans and African Americans [45, 202]. Furthermore, the hypertensives and normotensives in this study were identified from the same semirural clinics. The normotensive subjects were younger in mean age than the hypertensives but were recruited from decades in which hypertension is manifest. Since hypertension has a variable age of onset, it remains possible that some of the normotensives may subsequently develop hypertension, but this would only serve to dilute any association with this trait. Although the numbers of each sex were not equal in each group, it is unlikely that there is a sex-specific influence on the distribution of this intragenic variant of the β2 adrenoceptor, since the gene is located on chromosome 5.
Evidence for the regulatory influence of the Arg16→Gly variant of the β2 adrenergic receptor gene stems from transfection experiments in Chinese hamster (CHW-1102) fibroblasts [197], which indicate identical affinities of either variant for isoprenaline and adrenaline. In contrast, after prolonged exposure to isoprenaline, the Gly16 allele exhibited enhanced agonist-mediated receptor downregulation [197]. In collaboration with the group of M. Hughes (Hammersmith Hospital, London) we were able to provide first evidence in humans, that the Gly16 variant may be prone to exaggerated agonist-mediated downregulation [203], as evidenced by a reduced Bmax measured by PET scan (in vivo assessments of pulmonary β-adrenoceptor density after exposure to the β2-agonist salbutamol). Indeed, the Gly16 allele has been recently associated with nocturnal asthma [204], and therefore, diminished receptor number may be an important genetic factor in that asthmatic phenotype.
As far as cardiovascular regulation in subjects with β2 adrenergic receptor variants is concerned, we were able to show in a recent study, that the vascular response to infusion of the β2 agonist salbutamol may be modulated by the receptor genotype [205], so that subjects who are homozygous with respect to the pro-downregulatory Gly16 variant of the β2 adrenoceptor show a reduced fall of the total peripheral resistance index (Δ TPRI -17.9 ± 14.4 versus -30.6 ± 8.3%, p=.01).
In addition to differences on vascular responses to infused β2 adrenoceptor agonists, basal mean arterial blood pressure average (bMAPa) appears to be lowest in Arg16 homozygotes and highest in Gly16 homozygotes [205] (75.2 ± 4.93 versus 81.6 ± 6.14 mmHg, p < .01), respectively (Figure 5.1).
|
|
Additional support for a role of the β2 adrenergic receptor gene in hypertension arises from a recent report of association of a restriction fragment length polymorphism (RFLP) at this locus with hypertension in African Americans [193]. Although the Arg16→Gly variant, which may have functional influences, was not studied by that group, it is known that the used BanI restriction site is located at the position of the A1515G mutation, which is in linkage disequilibrium with the Arg16Gly coding alteration.
The distribution of the Arg16Gly β2 adrenergic receptor alleles in this study deviates from Hardy-Weinberg equilibrium. When deviations from this equilibrium arise, the possibility of selection bias in establishing the study or incorrectly assigned genotypes due to laboratory error must be considered. As we indicated, previous studies in this population have conformed to Hardy-Weinberg criteria, and we have genotyped all subjects twice, which makes these possible explanations less likely [45, 165]. The second analysed mutation (Gln27Glu), however, did conform to Hardy-Weinberg equilibrium. Interestingly, the distribution of RFLP genotypes of the β2 adrenoceptor reported previously also deviates from Hardy-Weinberg equilibrium in a population of African Americans [193]. Furthermore, in the same report from the same population, the genotypic distribution at the α2 adrenoceptor locus did confirm to Hardy-Weinberg criteria. A recent study about genetic variations of the β2 adrenergic receptor gene in Caucasian twins confirmed excessive homozygosity for the Arg16Gly mutation [206]. In tandem with our data, this finding may reflect the relationship of genotypes at the β2 adrenoceptor locus with hypertension rather than population bias or laboratory error.
This study offered 95% power to demonstrate a change in the distribution of the glycine allele from 0.64 in the normotensives to 0.85 in the hypertensives (χ2= 18.9, p = .0002). In spite of the power of this study and the observations reported, it is necessary to remain cautious about these results until they are replicated in other populations of the West African ancestry.
In conclusion, essential hypertension in African Caribbeans is associated with an increased frequency of the Gly16 allele of the β2 adrenoceptor. This variant may predispose to essential hypertension by conferring enhanced agonist-mediated receptor downregulation and represents an additional candidate for the genetic basis of this complex trait.
The Caerphilly Study. This study investigated four SNPs of the human β2 adrenergic receptor as candidate molecular variants predisposing to essential hypertension. The important findings of this study were that three of the four analysed mutations (5’ LC Arg19Cys, C1244T, and Glu27Gln) were associated with blood pressure. There was no association present with any examined quantitative traits and the Arg16Gly variant in this study. The significant associations we observed were for pulse pressure and systolic, not for diastolic blood pressure. This finding should not be surprising, because earlier studies indicate that systolic and diastolic blood pressure may be influenced by different genes [207].
Evidence for the regulatory influence of the Gln27→Glu variant of the β2 adrenergic receptor gene stems from transfection experiments in Chinese hamster (CHW-1102) fibroblasts [200], which normally do not express β2 adrenoceptor. The Glu27 variant displayed very little agonist-promoted downregulation, but agonist binding and coupling to adenyl cyclase were maintained.
A modulation of forearm blood flow by β2 adrenoceptor variants has been shown recently [208]. Subjects homozygous for the Gln27 receptor genotype had lower baseline blood flow compared with the Glu27 homozygotes (3.21±0.34 vs. 4.43±0.64 ml/100ml/min; p=0.05). The forearm blood flow response to isoprenaline was also attenuated in the Gln 27 homozygotes at all doses. Blood flow increased to 13.95±1.53 ml/100ml/min at the highest dose of isoprenaline (300 ng/min) in subjects with the Gln 27 genotype compared with 19.94±3.48 ml/100 ml/min in the Glu 27 homozygotes (p<0.05). Thus there is a relationship between the Gln27→Glu β2 adrenoceptor polymorphism and forearm vascular activity to isoprenaline in a group of male normotensive subjects. A physiological role for this polymorphism also has been shown in studies on airway responsiveness. Asthmatic subjects homozygous for the Glu27 genotype have less reactive airways than the Gln27 homozygotes, suggesting that the β2 adrenergic receptor polymorphism may be important in determining airway responses [209].
Attenuated vasodilatory response in people with the Gln27 variant might explain the strong association of this genotype with higher systolic blood pressure in the Caerphilly Prospective Heart Study. The 5’ LC Cys19 variant, which shows reduced receptor expression [171], also shows strong association with higher systolic blood pressure and pulse pressure. Interestingly, this variant is in strong linkage disequilibrium with the Gln27 variant (Table 4.10), making both of these changes work in tandem on blood pressure regulation.
|
It has to be emphasised that allele distribution of the Arg16Gly and the Gln27Glu variant as well as the observed linkage disequilibrium differed markedly between the two ethnic groups investigated in this thesis (Table 5.3). The Gly16 allele was more frequent (67%) in normotensive African Caribbeans than in Caerphilly individuals (46%, χ2=8.52, p=0.0035). The Glu27 allele was significantly more frequent in the Caerphilly data set (31%) than in the normotensive African Caribbeans individuals (16%, χ2=13.30, p=0.0003). These findings match nicely the data from other ethnic studies [210, 211, 212]. As the frequency of the receptor downregulation increasing variant Gln27 is high but uniform in the studied African Caribbeans (Table 3.4), this effect might explain the impaired vasodilation in people of West African ancestry, which has been reported in African-Americans [201]. We have no data on linkage disequilibrium between the 5’ LC Arg19Cys polymorphism and the Gln27Glu variant in African Caribbeans subjects so far, so there might also be speculated about the influence of the promoter polymorphism for blood pressure regulation in this ethnicity.
| |||||||||||||||||||||||||||||||||||||
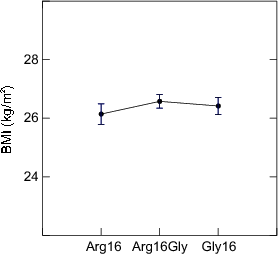
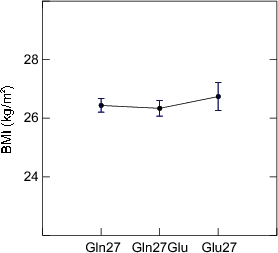
The close relationship between body mass index and blood pressure [213] is one of the most consistently observed associations between cardiovascular risk factors. The β2 adrenoceptor is a major lipolytic receptor in human fat cells [214]. The Glu27 polymorphism has been associated with higher BMI in a Swedish population (BMI of 30.7±1.5 for Gln27 homozygous versus 38.2±1.8 kg/m2, p=0.004 for Glu27 homozygous individuals) [215]. Interestingly, a similar study in a French population [216] showed men bearing the Gln27 genotype had higher body weight, BMI, and waist to hip circumferences. In the Caerphilly data set, we were not able to find any association between the codon 16 or 27 polymorphisms and body weight or BMI (Figure 5.2). Age and BMI were, nevertheless, strong predictors for systolic blood pressure and therefore regarded as important covariates in the ANOVA analyses.
|
Multiple linear regression analyses in both phases resulted in the β2 adrenoceptor genotype to have significant impact on blood pressure. Age and BMI were found to be important covariates. Interestingly, alcohol consumption was a significantly confounding variable in systolic blood pressure and pulse pressure only, whereas smoking was a confounding variable in diastolic blood pressure only. Alcohol consumption was positively correlated with systolic blood pressure, but not with diastolic blood pressure. Although several studies correlate alcohol consumption with increased systolic and diastolic blood pressure [217, 218], a recent study on Australian males showed similar findings. Alcohol consumption was significantly related to systolic (p=0.05) but not diastolic (p=0.25) blood pressure [219].
Although it is known that blood pressure and heart rate increase during smoking, several epidemiological studies have found lower average blood pressure values in smokers than in non-smokers [220]. Most studies are in accordance with our findings, that smoking is inversely related with diastolic blood pressure only [221, 222]. Interestingly, a qualitative variable coding for cardiovascular events in first degree relatives before the age of 55 emerged as significantly confounding variable for both, diastolic blood pressure and pulse pressure. This was not the case for cardiovascular events of first degree relatives without limiting for age.
The 5’ part of the β2 adrenergic receptor mRNA is a highly conserved structure. Nucleotide sequences of the 5’ LC are 70% conserved between species (Figure 5.3). The human sequence used as reference is based on the first cloned β2 receptor gene [111]. Interestingly, the other mammalian sequences in Figure 5.3 all show the Arg allele (CGC) at position 19 whereas the frequency for the Arg19 allele in the examined Caerphilly population was only 28%. The Arg19 allele shows higher receptor densities [171] and significantly lower systolic blood pressure in the Caerphilly data set.
|
|
The 5’ start of the β2 receptor gene is highly conserved as well (67%, Figure 5.4). Allelic distribution (Table 5.2) for the Arg16Gly and Gln27Glu polymorphisms suggests the Gly16 variant (54%) as well as the Gln27 (69%) variant to be termed wild-type. The first cloned human β2 adrenergic receptor sequence [111], usually referred to as reference sequence or wild-type, is Arg16 and Gln27. Alignment of seven mammalian species shows that these two positions are Gly16 and Glu27 in all compared mammalian sequences, which is concordant with the allele distribution in the Caerphilly data set for position 16, but differs significantly for the position 27 variant (Glu27 31%).
|
Sus 1752 T--GCGCCATGGGGCAGCCCGGGAACCGCAGCGTCTTCTTGCTGGCGCCCAACGGA Felis catus 1 GCGCCATGGGGCAGCCCGGGAACCGCAGCGTCTTCTTGCTGGCGCCCAACGGA Hamster 203 TGCCAGCGATGGGGCCACCCGGGAACGACAGTGACTTCTTGCTGACAACCAACGGA Mouse 2204 TGCCAGCCATGGGGCCACACGGGAACGACAGCGACTTCTTGCTGGCACCCAACGGA Rat 2249 GCCATGGAGCCACACGGGAATGACAGCGACTTCTTGCTGGCACCCAATGGA Macaca 33 TGCGCGCCATGGGGCAACCCGGGAACGGCAGCGCCTTCTTGCTGGCACCCAACGGA Human 1256 TGCGCGCCATGGGGCAACCCGGGAACGGCAGCGCCTTCTTGCTGGCACCCAATAGA Common TGCG.GCCATGG.GC..C.CGGGAA...CAG.G.CTTCTTGCTG.C..CCAA..GA 1 MetGly......GlyAsn...Ser...PheLeuLeu......Asn... Sus 1808 AGCCATGCGCCGGACCAGGACGTCCCACAGGAGCGGGACGAGGCATGGGTGGTGGGC Felis catus 54 AGCCACGCGCCGGACCAGGACGGCACGCAGGAACGAAACGACGCGTGGGTGGTGGGC Hamster 259 AGCCATGTGCCAGACCACGATGTCACTGAGGAACGGGACGAAGCATGGGTGGTAGGC Mouse 2260 AGCCGAGCGCCACACCACGACGTCACTCAGGAACGGGACGAAGCGTGGGTTGTGGGC Rat 2299 AGCCGAGCGCCAGGCCACGACATCACTCAGGAACGGGACGAAGCGTGGGTGGTGGGC Macaca 89 AGCCATGCGCCGGACCACGATGTCACGCAGGAACGGGACGAGGCGTGGGTGGTGGGC Human 1312 AGCCATGCGCCGGACCACGACGTCACGCAGCAAAGGGACGAGGTGTGGGTGGTGGGC Common AGCC..G.GCC...CCA.GA...C.C..AG.A..G..ACGA.G..TGGGT.GT.GGC 17 Ser......Pro......Asp............Arg.........TrpValValGly
|
The impact of the Gln27 downregulatory effect was obviously greater in the analysed Caerphilly data set than that of the Gly16 allele. Green et al [197] reported that cells transfected with the Arg→Gly16 substitution underwent a greater degree of downregulation (41 ± 3%, p < 0.05) than the Arg16 variant (26 ± 3%). Although the Gln27→Glu27 mutation displayed no downregulation whatsoever, the receptor with both mutations (Gly16 + Glu27) also displayed a greater degree of downregulation as compared to that found with the Arg16/Gln27 wild-type (39.0 ± 3.5%, p < 0.05). In our study, the Gly16 + Glu27 subjects had displayed a significantly lower mean systolic blood pressure than Gly16 + Gln27 subjects (Table 5.5). This surprising finding might be explained when taking into account that the experiments of Green were done using the originally cloned cDNA [111] subcloned into pcDNA 1/Neo using the expression vector’s own promoter whereas haplotype analysis (Table 4.10) showed that in our sample set the Glu27 variant was always associated with the 5’ LC-Arg19 variant. This makes it possible that the in vivo effect might well be more a 5’ LC-Arg19Cys receptor expression alteration than the changed receptor downregulation by Gln27Glu.
| |||||||||||||||||||||||||||||||||
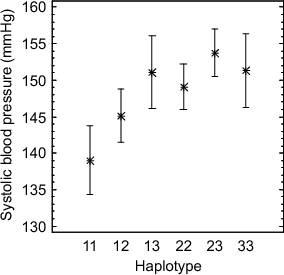
Haplotype analysis of the Caerphilly data set resulted in 3 main haplotypes, type 1 (5’ LC-Arg19 1242C Gly16 Glu27), type 2 (5’ LC-Cys19 1242T Arg16 Gln27), and type 3 (5’ LC-Cys19 1242T Gly16 Gln27). Type 3 (harbouring the receptor expression reducing 5’ LC-Cys19, prodownregulatory Gly16 and downregulating Gln27 variants) showed higher mean blood pressure than type 2 (which exchanges the pro-downregulatory Gly16 with Arg16). Interestingly, the heterozygous subjects (type 23) showed higher mean blood pressure than the homozygotes (type 33) in phase II, but not in phase III (n.s., Figure 4.14). Genotypes that comprise of one completely downregulating allele only (type 13) show the same effect on blood pressure than type 33 homozygotes (Figure 5.5) in both phases. It should be noted, that the 5’ LC-Arg19-Gly16-Glu27 homozygotes (type 11) show the lowest mean blood pressure in both phases, significantly lower systolic blood pressure than genotypes 13, 22, 23, and 33. Interestingly, the 11 genotype is the genotype present in all compared mammalian species (Figure 5.3 and Figure 5.4).
This study establishes an association between the 5’ LC Arg19Cys, C1244T, and the Gln27Glu polymorphism of the human β2 adrenergic receptor gene and essential hypertension. The β2 adrenoceptor genotype is significantly associated with systolic blood pressure as well as pulse pressure. We cannot state for certain which polymorphism we examined is responsible for the association with systolic, diastolic blood pressure and pulse pressure, nor can we speculate wether or not the functional polymorphisms exert independent effects. This may be elucidated by larger studies with complex haplotype analyses. Nevertheless, this study shows that at least 2 functionally relevant β2 adrenoceptor polymorphisms are associated with blood and pulse pressure. Finally, our study suggests that the 5’ LC-Arg19 + Glu27 variant is associated with lower blood pressure and a lesser risk to develop hypertension in white subjects.
|
|
© 2001 Alexander Binder