|
|
|
|
|
![]()
Extensive studies on the molecular features of the b-adrenoceptors have been carried out and the b-adrenoceptor is now among the best defined of the G-protein-linked superfamily of receptors.
The human b-adrenoceptors (b1, b2 and b3) are derived from at least three distinct genes and comprise of 477, 413 and 408 amino acids respectively. All three b-adrenoceptors show high amino acid sequence homology for each receptor between species. For example the b1 receptor of the mouse and rat are 90% homologous to the human b1. Similarly the mouse b2 receptor is 95% homologous to the human b2. However there are marked sequence differences between the subtypes. The b1 shows 48.9% homology with b2 and 50.7% homology with b3, whereas b2 show 45.5% homology with the b3.
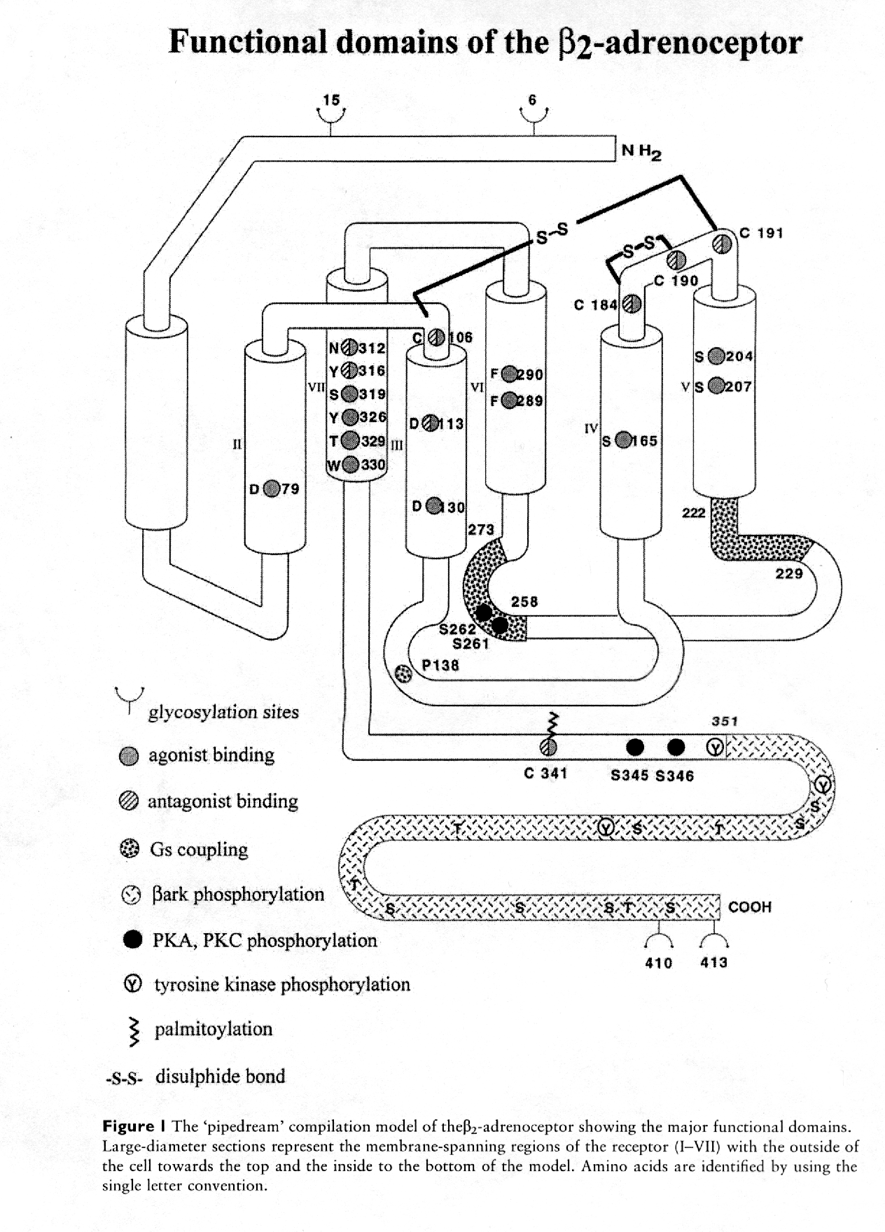
The G-protein superfamilies are characterised by their seven hydrophobic transmembrane spanning domains. Three extracellular and three cytoplasmic (intracellular) loops, with a glycosylated extracellular amine (N) terminus and cytoplasmic carboxy (C) terminus link the transmembrane domains. The seven transmembrane domains arrange in the membrane to form a "pocket".
A great deal of the research done to determine the functional domains of the b-adrenoceptors has been carried out on the b2 receptor. Many of these studies have involved the preparation and expression of site directed mutations (deletion or substitution) of particular amino acids and also the synthesis of chimeric receptors.
Diagram from Jrnl of Autonomic Pharmacology 17, 331-343.
Mutation experiments revealed that ligand binding appeared to occur within the transmembrane pocket. Fluorescently labelled b antagonist (e.g. carbazolol) confirmed this demonstrating that the location of the binding site was approximately 10.9A from the surface of the cell located within the membrane bilayer. Evidence in support of this also came from the deletion of the amino and the carboxy termini, which proved to have no effect on ligand binding.
Important residues involved in ligand binding also include Asp79, 113, and 130. Asp79 appears to be essential for the binding of high affinity agonist but is not required for the binding of antagonists. Asp113 located in the third transmembrane domain is vital for binding of both agonist and antagonist in the b2 receptor. Substitution of this residue with a Glu not only results in a marked decrease in the ability of the receptor to couple to adenylate cyclase but also appears to converts b antagonists to partial agonists and increases the efficacy of partial agonists. Substitution of Asp130 increases the affinity of the receptor for the agonist but reduces the ability of the receptor to couple to the G-protein.
Other important amino acids associated with binding include Ser204, 207 & 309; Tyr326 and Phe289, 290 alteration of these residues by substitution resulted in an increased affinity for agonist with no effect on antagonist binding. Ser204 forms hydrogen bonds with the meta hydroxy group of catecholamines; similarly Ser207 interacts with the para hydroxy group group of catecholamines.
Chimeric receptor studies revealed that transmembrane domains six and seven were important in determining the specificity of antagonist binding.
Residues of the third intracellular loop are involved in the coupling of the receptor to the Gs-protein, where the deletion of amino acids 222-229 and 239-272 caused complete loss of coupling to adenylate cyclase furthermore deletion of residues 258-270 reduced coupling. Alterations of the carboxy terminus to position 345 had no marked effect on receptor coupling however substitution of Cys341 the palmitoylation site with a Gly resulted in a non-palmitoylated receptor with reduced Gs coupling. Deletions of the carboxy terminal region of the third intracellular loop also resulted in a reduction in Gs coupling. Deletions in intracellular loop one resulted in impaired expression of the receptor. Deletions in loop two had very little effect however substitution of Asp130 for Gly reduced Gs coupling.
All of the b adrenoceptors have a glycosylated extracellular site on the N terminus; these sites are not associated with binding or signal transduction however it is thought that they are important for insertion of the receptor into the membrane. b adrenoceptors also contain a number of Cys residues (Cys106, 184, 190 & 191) which are thought to form disulphide bonds between the hydrophilic extracellular loops, which stabilise the receptor conformation.